
Les bactéries sont partout autour de nous, mais aussi dans notre intérieur (poumons, intestins …) . La biologie pastorienne les a identifiées, pour mieux les contrôler sur un plan médical, mieux les asservir pour des processus bio-industriels. Mais après un siècle de belligérance, on s’aperçoit que “l’adversaire” est le plus souvent un partenaire, et qu’il existe dans son milieu naturel, de manière totalement différente que dans nos cultures de laboratoires. Avec de telles erreurs de jugement, il n’est pas étonnant d’assister à des échecs thérapeutiques ou technologiques, en particulier lorsque les bactéries s’organisent en commun pour créer des biofilms.

Leur satisfaction fait plaisir à voir: “ils” ont décodé le génome de “la” bactérie Echerichia x, ou Bacillus y, c’est un grand pas en avant pour soigner telle affection ou pour dépolluer tel sol “. Parfait! Toutes les semaines, on a droit au même chant d’allégresse … qui n’est qu’un refrain. Car c’est bien d’avoir la carte mère d’un ordinateur, c’est mieux de savoir le brancher en réseau pour en utiliser toutes les ressources.
Les bactéries, si on les dissèque, sont des organismes effectivement très simples. Qui ont été étudiés et classifiés selon les mêmes critères que les angiospermes ou les primates: une morphologie, une spécificité.
Un travail de bénédictin, dans des laboratoires où l’on a utilisé des millions de boites de Pétri pour, repiquage après repiquage, isoler des souches pures, puis comprendre le fonctionnement ( nutrition, déplacement, métabolisme, reproduction) de chaque espèce.
Sur ce catalogue très fourni, mais purement descriptif, on a rajouté toutes les données de la génétique, on a pu alors découvrir que des centaines de gènes étaient partagés entre ces bactéries, des virus , et … nous mêmes. Et ce travail fondamental a été très fructueux, on a en effet grâce à ce modèle biologique très simple, à la reproduction ultra-rapide, compris des pans entiers du fonctionnement d’enzymes et de la chaîne ADN / protéines, des relations avec les virus, avec les cellules de métazoaires.
C’est que “la” bactérie telle qu’on la concevait, ne fonctionne pas toute seule avec un environnement de type pH / nutriments / température, mais dans un contexte beaucoup plus vaste et complexe, qu’on pourrait simplifier en observant autre chose, en changeant la focale du microscope.
A partir d’une conduite le nez dans le guidon, on est mieux inspiré de regarder le paysage … et apprécier le concept de diversité.
Diversité ! C’est le maître mot des écologistes, depuis des dizaines d’années. Qui n’est pas entré dans des oreilles formatées par les valeurs de production, d’individualité, de croissance.Et pourtant, il va falloir intégrer ce mot à tous les niveaux de compréhension des phénomènes biologiques complexes.
Le mythe des bactéries unicellulaires
Depuis qu’on a su cultiver des bactéries, qu’on a mis au point les supports nutritifs et des appareils d’observation performants, on a travaillé selon un dogme intangible, selon lequel lorsqu’on isole une bactérie et que l’on met en route sa multiplication, dans un milieu homogène donné, on obtient des millions de bactéries à l’identique, même contenu, mêmes physiologies.
En fait, dans ces conditions idéales de laboratoire, les bactéries en culture ne manquent de rien, et toute leur activité se résume à fabriquer une autre bactérie. Chaque individu consomme des nutriments et synthétise éléments de membranes, de parois et de cytoplasme, jusqu’à doubler de taille. Et à ce stade, la bactérie se scinde en deux individus: la bactérie ne grossit pas, c’est la population bactérienne qui s’accroît…
A ce stade de croissance exponentielle, on peut penser que la bactérie est un organisme immortel, et que chaque individu peut se reproduire à l’infini (reproduire dans l’esprit du clonage, et non dans l’esprit de la reproduction sexuée).
A la vitesse de croissance de la colonie, le milieu va immanquablement se trouver en déficit d’un nutriment, ce qu’on appelle un facteur limitant. La croissance se trouve alors limitée (phase de ralentissement), avec corrélativement une modification du métabolisme des bactéries.Il existe en effet une plasticité, une capacité pour les cascades enzymatiques, de contourner un problème, de solutionner une carence.
Car dans son historique de plusieurs millions d’années, cette bactérie a déja tatonné, elle a déja été confrontée à cette déficience, et elle a mis de coté (dans son génome) de quoi repartir du bon pied.
Et cette variation de métabolisme va entrainer la synthèse de métabolites secondaires, des produits (des déchets?) qui n’étaient pas produits jusqu’ici en phase exponentielle. Et qui vont peu à peu s’accumuler dans le milieu pour le modifier encore plus. Pas seulement, il y a souvent aussi des enzymes.. .
Ce remaniement du milieu devient progressivement fatal à certaines bactéries dont la division devient laborieuse, jusqu’à donner des bactéries filles incomplètes, non viables.
C’est le début d’une phase de déclin qui modifie totalement, et la population bactérienne, et le milieu (ce qui est totalement lié …). Le milieu devient hétérogène, et fluctue avec le temps.
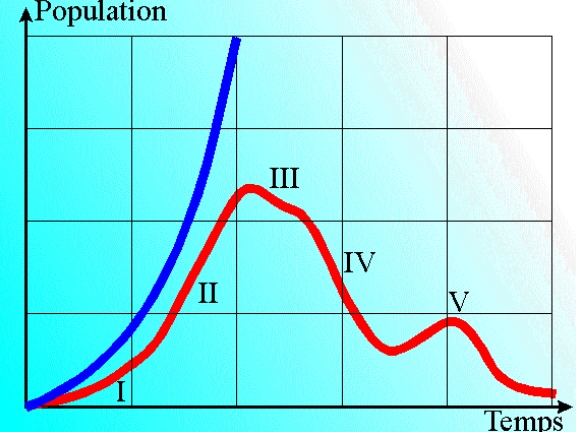
Quant aux bactéries, certaines meurent , se lysent et leur contenu vient encore modifier le milieu, d’autres modifient leur apparence par l’apparition d’organelles de fixation ou au contraire de mobilité (pili, flagelles), d’autres enfin “préfèrent” attendre des temps meilleurs par la sporulation, un enkystement autonome qui protège le génome et un concentré d’enzymes essentiels avec une résistance considérable à tous les stress, y compris la chaleur.
Au total, à partir d’une souche (à la limite, ce pourrait être à partir d’un seul individu) bactérienne, qui au début était intangible, on obtient tout naturellement une population d’individus qui portent le même nom, mais qui ont des réactivités biologiques toutes différentes.
Voici ce qu’on observe en laboratoire, dès qu’on s’écarte des conditions idéales de la croissance exponentielle. Mais dans la Nature, où tout varie, le jour, la nuit, les saisons, l’hygrométrie … les modifications de milieu sont nettement plus importantes et tous nos écosystèmes comprennent donc des communautés bactériennes dont certaines sont (momentanément) en phase exponentielle, d’autres en phase de ralentissement ou de déclin. Un joyeux mélange qui justement constitue la vie . Ou la mort de certains.
Un petit apparte au passage, qui concerne les humains: depuis deux siècles, l’homme s’est comporté comme ces bactéries en phase exponentielle, à qui il ne manque rien : eau, oxygène, ressources énergétiques et alimentaires. D’où une expansion démographique couplée à un bien-être qui semblent sans limites : c’est le mythe du Progrès. Or, on s’aperçoit que certaines ressources sont irrémédiablement entamées ou détruites (eau, poissons, humus, pétrole) et que le milieu est déja passablement dégradé. Et tout ça, comme nous le chante Cabrel, “dans le même aquarium”.Nous sommes donc en phase de ralentissement. A gérer au plus vite.
Fin de l’apparte.
Ainsi donc, les bactéries possèdent en elles des capacités de modifier leur métabolisme, au moindre écart dans les conditions de milieu. Les cultures pures, celles qu’on réalise par exemple pour faire un antibiogramme ou pour “faire pousser” les bactéries d’un prélèvement d’urine, sont des artéfacts de microbiologistes (je cite Janine Guespin-Michel), et il n’est pas étonnant que des remèdes qui semblent juguler une colonie en boite de Pietri, soient inefficaces (pas toujours, tout de même!) lorsque appliqués au malade …
Dans la nature, les bactéries vivent en communautés hétérogènes, déja du fait des fluctuations du milieu, mais également parce qu’il est rare d’avoir en fait une seule espèce bactérienne en un lieu.
Cette hétérogénéité est un désastre médical dans certaines situations nosocomiales, mais c’est un atout précieux après une catastrophe environnementale de type marée noire.
Les recherches en bactériologie se sont déroulées en “support mou”, au fond de boites de Pétri, où les bactéries se multiplient sans obstacle en trois dimensions. Mais dans la nature, les bactéries sont sans cesse sollicitées par des supports, ce qui va orienter leur développement.
Ainsi, avec des substrats en suspension, comme des cristaux ou de la fibrine dans une urine pathologique, les bactéries vont développer des colonies.
Sur des supports physiques rigides, les bactéries vont d’abord coloniser la surface avec des productions gluantes, avant de s’y développer en compagnie d’autres organismes (bactéries, algues, flagellés, mycoplasmes) qui profitent ensemble de l’aubaine : c’est la création d’un biofilm.

Ainsi, on a selon le support deux types de développement: en milieu semi-solide, des bactéries de même type, apparemment semblables, mais déjà physiologiquement différentes, des clones presque parfaits, qui se multiplient à vitesse constante dans des directions concentriques en maximisant le nombre de contacts entre elles, avec des aspects géométriques caractéristiques de “la” bactérie et de son milieu.
Les laborantins exercés ne s’y trompent pas, et savent au coup d’oeil (et parfois au nez!), repérer et identifier une culture.
Dans les biofilms (voir illustration), aucune construction structurée. Les bactéries laissent apparaître des organites exocellulaires qui en modifient totalement l’aspect, et il n’y a aucune “géométrie” pour servir de repère à l’observateur.
Au contraire, ces bactéries glabres et rondelettes qui deviennent hirsutes et torturées, constituent des casse-tête , voire des pièges pour les laborantins qui doivent les discerner. L’aspect général d’un biofilm est celui d’un organisme multicellulaire, comme une sorte d’éponge, dont la matrice gluante est produite par l’ensemble des participants, bactéries, mais aussi monocellulaires, voire champignons …
Cette matrice est très hétérogène, elle nourrit différemment des bactéries de la même espèce en fonction des productions locales des autres habitants du lieu.
Cette même matrice constitue un obstacle pour des substances qu’on voudrait mettre en contact avec ces bactéries. Un obstacle primordial quand on veut désinfecter une gencive, utiliser un cicatrisant dans une plaie, ou permettre l’application impeccable d’une lentille de contact …
Cette matrice sans cesse renouvelée et qui en protège les locataires, on peut la décaper avec des tensio-actifs biologiques, ou la lyser avec des enzymes adéquats. Encore faut-il respecter à la fois le substrat biologique, et le voisinage cellulaire ou bactérien …
Encore un aparte: une gangue de polysaccharides qui protège une cible thérapeutique, ça ne vous dit rien ? C’est précisément le type de fourreau que les tumeurs édifient sur leur pourtour pour échapper tant aux cellules de l’immunité cellulaire, qu’aux médicaments de chimiothérapie locale.
Et l’analogie peut aller plus loin: ces cellules tumorales qui se multiplient à grande vitesse tant qu’il y a des nutriments, on les considère comme des clones, qui seront sensibles à un médicament unique qui aura fait ses preuves sur la cellule originelle.
Mais là encore, la matrice de ces cellules est sans cesse changeante, subissant des actions permanentes de l’organisme, des cellules tumorales elles-mêmes, et des thérapies anti-cancéreuses.
Comme les bactéries de notre sujet, les cellules cancéreuses ne peuvent pas rester du même type, certaines s’écartent du fonctionnement initial. Et ceci sans aller chercher des mutations chromosomiques ( c’est vrai qu’elles existent, mais à quel niveau?), simplement en laissant s’exprimer des gènes jusque là inactifs, mais que des modifications du milieu mettent manifestement en route
. Fin de l’aparte
Ainsi, lorsque le biologiste, mais aussi le médecin, doivent appréhender telle bactérie, quelles lunettes doit-il porter ? Un microscope pour zoomer sur le micro-organisme individuel, à comparer avec d’autres bactéries bien classifiées, une binoculaire pour observer un groupe homogène, mais dont on sait que la variabilité va se mettre en route, ou bien des lunettes de vue pour observer l’allure de cette communauté ?
Les trois, mon Capitaine,et plusieurs fois par jour …
Pour chaque bactérie-type, dix bactériophages régulateurs
Superbement gnorés par les labos car non brevetables (encore que …), les bactériophages seront obligatoirement des remèdes hospitaliers dans les années à venir.
Ces bactériophages sont surtout connus comme modèles d’étude, à la fois des virus et des bactéries: ils ont permis de bien comprendre les va-et-vient de gènes dans une souche de bactéries, la mise en route de fonctions éteintes (comme la virulence d’un Clostridium), ou au contraire le tarissement d’une production, dans des fermenteurs de l’industrie agro-alimentaire.
Si on estime à 10 puissance 30 le nombre total de bactéries, on doit considérer que les phages sont à 10 uissance 31, soit dix types de phages pour une bactérie. Ce différentiel de 10 est énorme.
Il nous indique que les phages ne se mettent pas à dix contre un pour simplement endiguer ou contrer le développement des bactéries. Certes, il existe une course poursuite de mutations entre bactéries et bactériophages, de type glaive / cuirasse, où d’ailleurs les bactéries sortent toujours en tête.
Mais cette prolifération des phages, qu’on pourrait qualifier “d’inutile” , leur omniprésence dans l’environnement naturel des bactéries, montre plutôt que ces virus font partie d’un processus de coopération réciproque, qui avait jusqu’ici échappé aux chercheurs.
Comme le souligne Janine Guespin-Michel, “les voir simplement comme des parasites ou des prédateurs, c’est négliger les interactions multiples qu’ils entretiennent avec les bactéries. Une hypothèse concernant leur rôle pourrait être qu’en lysant les bactéries, , ils contribuent à la croissance en hétérotrophie des populations en environnements carencés”.
Autrement dit, c’est en sacrifiant certaines bactéries dont le développement devient prédominant (bactéries en phase exponentielle), dont le contenu se déverse alors dans le milieu , contribuant ainsi à le modifier en l’enrichissant, que les phages spécifiques d’un clone bactérien pourraient permettre permettent le développement des autres types de bactéries.
Les phages seraient ainsi les garants d’une diversité maintenue ( tout du moins en milieu naturel …).
Mais on peut encore aller plus loin: puisque il y a échange régulier de gènes entre phages et bactéries, et ceci dans le cadre d’un équilibre des populations, ne peut on pas penser que les phages jouent le rôle de réservoirs de gènes externalisés? Des gènes inutiles pour une vie purement végétative dans un milieu riche en nutriments, mais qui constitueraient une “boite à outils génique” disponible pour toute modification de milieu qui impose un changement de métabolisme, voire une dessication de survie.

Alors ces bactériophages, ennemis permanents d’une gente bactérienne aux abois, ou bien régulateurs biologiques des modifications de l’environnement pour entretenir la diversité au sein des populations bactériennes?
Il n’y a aucun antagonisme entre ces deux propositions. En particulier si l’on oublie les individus (bactéries, phages) pour considérer les fonctions, si l’on prend du recul et qu’on considère les choses globalement.
Un troisième aparte:
Dans l’organisme des mammifères, les lymphocytes B produisent des anticorps (immunoblobulines) à l’encontre d’organismes extérieurs jugés inopportuns, parfois envers des tissus de leur propre organisme, mais ceci dans un cadre bien délimité qui s’appelle la santé.
Si cette production s’accélère ou perdure, d’autres lymphocytes B vont mettre en route de manière très active, la synthèse de nouveaux anticorps dirigés contre les précédents, on les appelle les anticorps anti-idiotypiques: ils vont d’une part neutraliser les anticorps en surplus dans le sang et les tissus, et d’autre part tarir la production des lymphocytes qui s’étaient emballés.
Voilà un autre exemple d’une action biologique, avec aussi bien un rôle de censeur que de régulateur …
Actions de proximité, actions à distance
Comme on a pu décrire le “gène égoïste”qui ne code que pour des protéines nécessaires à sa survie, on peut évoquer la “bactérie égoïste” qui ne se multiplie que pour sa fratrie.
Mais cette bactérie égoïste est vite obligée de partager … En effet, elle n’est jamais seule, et ses propres déchets, les métabolites qu’elle rejette dans son entourage, en modifient la composition, et servent de ressource à une autre bactérie, qui elle même produira des rejets , etc …
Sans le vouloir, cette “bactérie égoïste” met en route une coopération, qu’on appellera symbiose si elle profite aux deux, parasitisme si seule la seconde en bénéficie.
Puis un troisième organisme viendra ce greffer, puis un quatrième … et c’est alors que le milieu aura totalement changé, on pourra parler d’écosystème.
En milieu liquide, les bactéries ne sont pas bloquées contre un substrat, et peuvent être brassées pour développer une action sur un grand volume, une action à distance.
C’est le cas dans les industries basées sur les fermentations ( brasseries), c’est le cas au naturel dans une mare d’eau, et plus particulièrement dans la panse des ruminants: ici cohabitent et se développent plusieurs types de bactéries, chacune ayant un rôle dans la digestion des végétaux, les amylolytiques pour les grains, cellulolytiques pour les feuilles, dans un équilibre en phase avec le régime alimentaire.
Que cette brebis prenne en cachette une grosse ration d’avoine, et se développe d’un coup ( phase exponentielle) toute une flore amylolytique qui va acidifier le contenu du rumen, modifier anisi gravement la microflore locale et entraîner une gastro-entérite aigue.
Et puis il faut aussi considérer que toutes ces bactéries, après avoir joué un rôle enzymatique, sont elles-mêmes entrainées depuis le “digesteur” ruminal, vers l’intestin où elles seront elles mêmes digérées, apportant le quota protéique de la ration. On pourrait avancer que les herbivores sont aussi des “bactériovores”.
Nous avons désormais compris que notre terre est un petit village, où tout s’entrecroise. Y compris les effets des bactéries.
Les compositions de l’air et de l’eau des océans sont fixes, car extrèmement régulées par des phénomènes complexes bourrés de rétroactions, mais on n’est pas à l’abri d’un dérapage écologique avec effet d’emballement.
Comme lorsque des cyanobactéries , il y a deux milliards d’années, ont “inventé” la photosynthèse et fait monter le taux d’oxygène de l’air, de quelques pourcents à nos 21 % actuels, ravageant ainsi toute une flore existante, mais faisant le nid de toutes les cellules actuelles équipées de mitochondries …
C’est ainsi que les chercheurs lorgnent actuellement sur certaines bactéries, en particulier celles qui entrainent la production de méthane, des bactéries stratégiques en cette période d’emballement climatique …
Ce petit tour d’horizon chez nos amies les bactéries, nous montre bien que de même que les physiologistes délaissent peu à peu la notion de tissu pour exploiter la notion de fonction (la neuro-immunologie, par exemple), nous devons considérer les bactéries comme des méta-organismes aux contours assez flous, mais dont on ne comprendra les mécanismes qu’avec un recul à plusieurs niveaux.
Un nouveau regard qui suppose de croiser les connaissances entre diverses disciplines. Ce qui ne correspond en rien au mode de fonctionnement de la recherche ( labos cloisonnés, revues savantes toujours plus pointues).
Jean-Yves Gauchet

Source: Un ouvrage passionnant a initié cet article: il s’agit de “Les bactéries, leur monde, et nous”, de Janine Guespin-Michel, professeure à l’université de Rouen. Un tableau des connaissances anciennes ou plus récentes qui permet d’apréhender les interactions réciproques et dynamiques, entre les bactéries, leur milieu immédiat, et les reste de notre monde. Collection Univescience – Co-édition La Recherche / Dunod